Глава 1. Рациональное питание беременных
Влияние количественных и качественных нарушений питания на течение беременности, развитие плода и состояние новорожденного (Е. П. Самборская, Б. В. Охрончук)
Значение полноценного питания женщины для нормального развития плода доказано многочисленными клиническими наблюдениями и экспериментальными исследованиями.
О связи питания матери с показателями физического развития плода и состояния его здоровья свидетельствуют и данные Л. Н. Антонова (1947), собранные в Ленинграде в период блокады города. Масса доношенных новорожденных в 1942 г. была в среднем на 550 г меньше, чем у новорожденных в 1940 г., а число детей, родившихся недоношенными, достигло 41,2%. Аналогичные данные об изменении показателей физического развития новорожденных в Чехословакии приведены Sabata (1963).
Согласно материалам ВОЗ (1971), средняя масса тела новорожденного в развивающихся странах ниже, чем в развитых странах. Это связано не с расовыми различиями, а с плохим питанием беременных женщин в развивающихся странах. Энергетическая ценность пищи, потребляемой женщинами Индии, родившими детей с массой 2500 г и ниже, составляла в последнем триместре беременности 6908 кДж (1650 ккал) в сутки (Shankae, 1962).
Недостаточное питание матери оказывает отрицательное влияние не только на массу плода, но и на его рост.
В экспериментальных исследованиях по изучению влияния голода и недостаточного питания на состояние самки и потомства установлено, что при неполноценном питании прерывается беременность, уменьшается численность помета снижается жизнеспособность и масса плодов (Payne, Wheeler, 1967; Smart и соавт., 1972).
Доказано, что нарушения питания в различные сроки беременности по-разному влияют на состояние плода (В. И. Бодяжина, 1962; А. Г. Пап и соавт., 1966). В экспериментах на мышах установлено, что голодание в предимплантационный период вызывает гибель зародышей, в период органогенеза - уродства развития, в фетальный период - физиологическую незрелость новорожденных (Мс Clure, 1959). Непродолжительное недостаточное питание может не отразиться на состоянии зародыша.
Для нормального развития плода и новорожденного важен не только количественный, но и качественный состав пищи. Имеются данные о влиянии на течение беременности, развитие плода и новорожденных нарушений качественного состава пищевого рациона беременных и, в частности, изменения содержания в нем основных пищевых веществ - белков, жиров, углеводов, витаминов, минеральных веществ и жидкости.
Белки являются основным пластическим материалом для построения тканей плода, синтеза гормонов и ферментов в организме матери. Учитывая незаменимость белков пищи другими пищевыми веществами, их дефицит существенно сказывается на развитии плода и состоянии беременной.
Наблюдения за беременными женщинами, проживающими в странах с низким экономическим уровнем, показали, что самопроизвольные аборты, преждевременные роды и повышенная перинатальная смертность чаще всего наблюдались у женщин малоимущих классов, употребляющих пищу, бедную белками и богатую углеводами (Thomson, 1959).
В эксперименте показано, что малобелковое питание снижает плодовитость животных. Это проявляется как уменьшением числа эффективных спариваний, так и снижением численности помета; выжившее потомство отличается слабой жизнеспособностью, отставанием в росте и развитии; в печени и мозге у родившихся животных снижено количество функционально полноценных клеток (Kohrs и соавт., 1976; Nancy и соавт., 1978).
Даже временный дефицит белков ведет к задержке развития плодов и снижению массы их тела, мозга, печени и сердца (Аrауа и соавт., 1978; Shinn, Goshiaki, 1978).
Недостаточное белковое питание приводит к изменению биохимического состава крови и мышцы матки, что способствует изменению ее реакции на беременность. С точки зрения Leathema и соавторов (1968), этим и объясняется нарушение репродуктивной функции при недостаточном белковом питании.
Имеет значение не только количество белков в рационе питания беременной, но и их качественный состав. Известно, например, что плод, используя при голодании матери ее белки, прежде всего усваивает глобулины и в меньшей степени альбумины, поэтому изменения соотношения альбуминов и глобулинов в сыворотке крови у беременной обычно нарушают эмбриональное развитие (Langman и соавт., 1959).
Согласно экспериментальным данным, недостаток лизина в рационе влияет на массу и рост крысят (Stapleton, Hill, 1972). При дефиците в рационе лизина или метионина затрудняется родовая деятельность, увеличивается смертность самок в родах и смертность потомства, снижается масса тела новорожденных (Stapleton, Hill, 1980).
Количественный и качественный состав жиров в рационе беременных весьма существенно влияет на потомство. В эксперименте доказано, что при недостатке жиров в питании уменьшается численность помета, масса тела новорожденных крысят и нарушается их постнатальное развитие. Уменьшается также масса мозга и содержание в нем холестерина, ДНК и РНК (Borgman и соавт 1979; Crawtord, Hassan, 1979). Изучая роль ненасыщенных жирных кислот, Crawtord, Hassan (1979) установили, что уменьшение их содержания в рационе беременных животных приводит к задержке становления рефлексов у крысят при нормальном развитии мозга. Длительное и значительное исключение из рациона беременных крыс жиров (за счет увеличения глюкозы и крахмала для сохранения энергетической ценности) приводит к снижению выживаемости потомства и падению массы тела крысят (Sinclair, Crawtord, 1973).
Избыточное поступление углеводов, особенно сахарозы в организм беременной значительно повышает частоту внутриутробной гибели плода (Kendall, 1961).
Известно, что глюкоза легко проникает через плацентарный барьер и поступает в организм плода (Glatzel, 1962), у беременных женщин с пониженной толерантностью к глюкозе чаще наблюдаются мертворождения чем у пациенток с нормальной способностью ее усваивать. У женщин с привычными выкидышами часто обнаруживается повышенное содержание сахара в крови и глюкозурия.
Витамины в пищевом рационе необходимы для нормального течения беременности, роста плода, подготовки. При недостатке витаминов возникают различные осложнения беременности (самопроизвольные выкидыши, преждевременные роды и др.). Во время беременности потребность в витаминах возрастает почти в два раза.
Потребность в витаминах группы В во время беременности повышена вследствие активации обменных процессов, в которых они принимают непосредственное участие.
Glatzel (1962) отмечает низкое содержание витаминов этой группы в форменных элементах крови у беременных женщин.
В эксперименте установлено, что недостаток витамина В1 (тиамина) в рационе крыс вызывает уродства конечностей у крысят, анэнцефалию и кровоизлияния в разные органы и ткани. Отмечается снижение плодовитости и выживаемости потомства (Pfeltz, 1955).
По данным Kelt (1959), исключение витамина В2 (рибофлавина) из рациона мышей с 9-го по 13-й день беременности вызывает у плода уродства развития головы, конечностей и органов пищеварительной системы.
Относительно влияния витамина В6 (пиридоксина) на беременность и внутриутробное развитие плода сведений в литературе мало.
По мнению Heller и соавторов (1973), гиповитаминоз В6 не оказывает существенного влияния на состояние беременной и развитие плода. Однако это противоречит имеющимся данным о роли витамина В6 в развитии ранних токсикозов беременных (И. И. Матусис, 1975). По данным Horalt (1954), дефицит витамина В6 приводит к неукротимой рвоте беременных, устраняемой введением пиридоксина. Hilman и соавторы (1962) отмечали противокариозное действие витамина В6 у беременных.
В то же время введение избыточных доз витамина В6 беременным мышам отрицательно влияет на развитие зародышей, вызывает нарушение белкового обмена (Morita, 1958).
Недостаточность витамина РР (никотиновой кислоты) может привести к аномалиям развития плода, прерыванию беременности, преждевременным родам и др. С дефицитом никотиновой кислоты связывают привычные выкидыши, мертворождения, снижение количества и качества молока у кормящей женщины (С. Н. Астахов, 1954).
При беременности может наблюдаться также дефицит витамина Н (холина). По данным Н. Дубнова (1952), отсутствие достаточного количества этого витамина в первые дни беременности вызывает изменения в плаценте и резорбцию эмбрионов.
Согласно данным Park, Vernoum (1973), при дефиците витамина В12 в рационе беременных крыс уменьшается масса тела новорожденных крысят; длина их существенно не изменяется. Отмечено также снижение активности ряда ферментов сыворотки крови животных и их устойчивости к гипоксии. С увеличением содержания витамина В12 в рационе масса тела новорожденных крысят увеличивается.
Недостаток фолиевой кислоты (фолацин, витамин Вс) при беременности у крыс приводит к лейкоцитопении, агранулоцитозу, ретикулоцитозу, уменьшению массы тела крысят, снижению аппетита и резорбции плодов. Избыток фолиевой кислоты подавляет синтез ДНК у зародышей лягушек (Grant, 1960).
Особое значение имеет обеспеченность организма беременной витамином С.
По данным большинства исследователей, с увеличением срока беременности возрастает потребность в витамине С (Е. И. Кватер и М. С. Маршак, 1962; Stojanow, 1963; Hiiter, 1965). Гиповитаминоз С является одной из причин преждевременных родов, выкидышей, рождения мертвых, с малой массой и неполноценных детей. В опыте на гвинейских свинках Stojanow (1963) установил, что кормление животных пищей с низким содержанием витамина С приводит к абортам, недонашиванию и летальным исходам. У умерших животных содержание витамина С в яичниках снижается до 85,2 мкмоль/л (1,5 мг%) и почти в два раза реже наступает зачатие.
Существуют различные мнения относительно влияния витамина С на родовую деятельность. По данным А. П. Николаева (1956), под влиянием витамина С происходит ускорение родов. В то же время Stojanow (1963) считает, что дополнительное введение витамина С увеличивает продолжительность родов, а его дефицит стимулирует родовую деятельность.
Имеются данные об активной роли плаценты в накоплении витамина С (Htiter, 1970); с увеличением срока беременности содержание витамина С в ней увеличивается.
Избыток аскорбиновой кислоты, по данным Е. П. Самборской (1964, 1966), приводит к прерыванию беременности, особенно на ранних стадиях развития зародыша. По мнению автора, абортивное действие витамина С связано с повышением уровня эстрогенов в организме и с изменением свертывающих свойств крови, приводящих к образованию ретроплацентарной гематомы с последующей отслойкой околоплодного яйца.
В эксперименте изучены нарушения, возникающие при недостаточности витамина А (ретинола) в пищевом рационе беременной. Доказано, что при этом у плода нарушается развитие сердца и кровеносных сосудов, возникает микроцефалия и анофтальмия (Sarma, 1959) При полном отсутствии витамина А в рационе с 14-го дня беременности в большинстве случаев возникает резорбция зародышей и гибель плодов. У оставшихся в живых новорожденных крысят скорость роста замедляется по сравнению с таковой у животных, получавших в рационе витамин А. При выраженном авитаминозе А вследствие гибели яйца становится невозможным зачатие (Joko и соавт., 1975).
По данным Gal и соавторов (1974), содержание витамина А и каротинов в сыворотке крови женщин с физиологическим течением беременности колеблется. Отмечаемый при беременности дефицит витамина А связывают с понижением способности печени в этот период превращать каротин в витамин A (Htiter, 1965).
Избыток ретинола для беременной так же вреден, как и его недостаток. В эксперименте на крысах при гипервитаминозе А у новорожденных наблюдались мозговые грыжи, анэнцефалия, патология мочеполовой системы, уродства развития глаз и конечностей (Giroud, 1957). Минимальная суточная доза ретинола, вызывающая тератогенное действие у крыс, по данным Cohlen (1953), составляет 2500 ЕД.
Витамин D (кальциферол), участвуя в обмене кальция и фосфора, способствует окостенению скелета плода и тем самым предупреждает возникновение рахита (М. М. Шехтман, 1976).
Передозировка витамина D приводит к раннему окостенению черепа плода (Е. М. Лукьянова, 1965; Huter, 1965) и снижению массы его тела (Lawistowski и соавт., 1970).
Витамин Е (токоферолы) оказывает влияние на течение беременности. В акушерской практике его применяют при привычных абортах, для лечения недонашивания и бесплодия. Он усиливает продукцию желтого тела или потенцирует его активность (М. М. Шехтман, 1966; Huter, 1965).
Установлено тератогенное действие дефицита витамина Е (А. П. Дыбан, 1959). В эксперименте на крысах показано, Что значительное увеличение в рационе беременных крыс витамина Е (до 500 мг/сут) ведет к аномалиям глаз у потомства.
Не менее важное значение имеет обеспечение организма беременной необходимым количеством макро- и микроэлементов. Избыток и недостаток их неблагоприятно влияет на организм беременной и плода. Поступление макро- и микроэлементов в необходимых количествах обеспечивает гомеостаз организма, способствует нормализации кислотно-основного состояния, активности ферментных систем и гормонов.
Кальций имеет важное значение для построения костно-хрящевых тканей плода. Установлено, что у беременной имеется дефицит кальция в связи с повышенной потребностью в нем плода (А. А. Покровский, 1966).
Huter (1965) считает, что наблюдающееся снижение содержания кальция у беременной связано не только с повышенной потребностью в нем плода, так как плод получает от матери до 30 мг кальция, то есть 3 - 4% резерва скелетного кальция, но также и с недостаточным содержанием его в рационе беременной. Если повышенная потребность кальция не покрывается содержанием его в пищевом рационе, то плод обеспечивает себя за счет материнского организма, что и приводит к его дефициту у беременной.
Фосфор так же, как и кальций, участвует в образовании костной ткани плода и необходим беременной женщине. Соотношение кальция и фосфора в рационе должно составлять 1:1,5; 1:2.
Имеются данные о депонировании фосфора в плаценте (А. С. Лесакова, 1957).
По наблюдениям П. Д. Давыдова (1962), прием в последние три месяца беременной препаратов кальция и фосфора способствует увеличению роста новорожденных в среднем на 1,5 см.
Нарушение развития плода происходит при дефиците и других минеральных веществ.
По данным R. S'poeri (1975), недостаток меди в пищевом рационе беременной приводит к внутриутробной гибели плода либо к рождению нежизнеспособных новорожденных с большими отеками, подкожными кровоизлияниями, грыжевыми выпячиваниями.
При дефиците цинка в рационе у беременных крыс наблюдается анорексия, в результате чего снижается поступление питательных веществ, возникает дефицит белка, уменьшается масса тела, селезенки, молочных желез и гематокритное число. При значительном дефиците цинка отмечены задержка роста и уменьшение массы плодов, а также гибель самок во время родов (Argon, 1976, 1977; Fosmire и соавт., 1977).
Железо имеет важное значение для беременной и для плода, так как входит в состав гемоглобина и участвует в окислительных процессах. Плод активно поглощает железо из организма матери, что создает его дефицит у беременной (Glatzel, 1962). Huber (1961), Schaumkell, Hollenbach (1961) обнаружили железодефицитную анемию в последние сроки беременности почти у всех обследованных женщин. Дефицит железа в рационе беременных белых крыс вызывает снижение массы плодов и анемию (Ahestrom и соавт., 1969).
Нередко отмечается недостаток нескольких минеральных веществ в пищевом рационе. Например, при недостатке йода часто наблюдается дефицит марганца. Недостаток йода и марганца понижает жизнеспособность новорожденных животных и вызывает у них отеки (Т. С. Самофал, 1964). Дефицит брома, молибдена, кобальта и йода приводит к уродствам развития (Е. М. Мещенко, В. И. Алексин, Л. И. Котелянская, 1959).
Питание, содержащее оптимальное количество макро- и микроэлементов, не только обеспечивает нормальное течение обменных процессов у матери и плода, но и является важным лечебным фактором (Broman, Malmstrom, 1963).
Наблюдения, проведенные А. Г. Коломийцевой и соавторами (1969), показали, что применение комплекса микроэлементов при позднем токсикозе беременных нормализует обменные процессы, препятствует развитию ряда осложнений.
Одной из особенностей водно-солевого обмена у беременных является тенденция к задержке жидкости в организме. Этому способствует накопление натрия хлорида в тканях и повышение функции передней доли гипофиза, сопровождающееся усилением выработки антидиуретического гормона. Во время беременности имеет место разжижение крови и повышение проницаемости кровеносных сосудов, что создает предпосылки для образования отеков.
Увеличение объема циркулирующей крови, в основном за счет плазмы, приводит к относительному снижению содержания гемоглобина и эритроцитов, а также белков плазмы (Л. С. Персианинов, В. Н. Демидов, 1972).
Приведенные данные свидетельствуют о значении полноценного питания беременной для нормального развития плода и новорожденного. Полноценными следует считать рационы, в которые входят белки, жиры, углеводы, минеральные вещества, витамины, жидкость в оптимальных для беременной количествах и соотношениях. Сбалансированным является такое соотношение пищевых веществ в рационе, которое может обеспечить нормальное функционирование всех систем и органов.
Поэтому представляется чрезвычайно важной разработка научно обоснованных рекомендаций по составлению сбалансированных рационов для беременных женщин, рожениц и родильниц.
В 1982 г. коллегией Министерства здравоохранения СССР утверждены новые нормы физиологических потребностей в пищевых веществах и энергии для различных групп населения СССР, в том числе и для беременных женщин (см. табл. 4).
В Киевском НИИ педиатрии, акушерства и гинекологии на основе проведенных клинико-экспериментальных исследований и данных литературы составлены и внедрены в областях республики научно обоснованные дифференцированные рационы питания для беременных женщин с учетом массы тела, роста, характера трудовой деятельности, энерготрат и срока беременности.
Разработке рационов предшествовали экспериментальные исследования по изучению влияния качественно различного питания на состояние беременных самок и их потомства и клиническое изучение фактического питания беременных женщин и роли его нарушения в возникновении осложнений беременности, родов и перинатальной патологии.
Экспериментальные исследования. В эксперименте на белых крысах линии Вистар с массой тела 180 - 200 г изучали влияние качественно различного питания на обменные процессы у беременных крыс, репродуктивную функцию самок, а также на показатели физического развития, состояния обменных процессов и красной крови у их плодов.
При составлении рационов с различным содержанием основных пищевых веществ использовали полноценный рацион вивария. Энергетическая ценность рационов для всех групп была одинаковой и соответствовала 339 кДж (81 ккал) в сутки. Применяли рационы с избыточным содержанием белка или избыточным содержанием углеводов. В зависимости от получаемого рациона животные были разделены на 4 группы. 1-я группа включала 60 небеременных белых крыс, находившихся на рационе вивария. Остальные группы включали беременных животных. Из них животные 2-й группы - контрольной (60 крыс) получали рацион вивария; 3-й (40 крыс) - рацион с избыточным (25% общей энергетической ценности) содержанием белка и соответственным уменьшением количества углеводов и 4-й (40 крыс) - рацион с избыточным содержанием углеводов (25% общей энергетической ценности), в основном, за счет крахмала и соответственным уменьшением количества белка.
Показатели белкового обмена. При анализе показателей белкового обмена установлено, что в динамике беременности у крыс всех групп выражена тенденция к снижению содержания общего белка, и уменьшается содержание альбуминов в сыворотке крови. У животных контрольной группы и у животных, находившихся на рационе с избытком углеводов, это снижение было более выраженным, чем у животных, получавших рацион с избытком белка. Количество альбуминов составляло 0,4285 ± 0,0075 от общего белка у крыс до зачатия; 0,4002 ± 0,0098 у беременных крыс контрольной группы, Р < 0,05; 0,4049 ± 0,009 - у беременных крыс, получавших рацион с избытком углеводов, Р < 0,05 и 0,4196 ± 0,0158, Р < 0,05 -у крыс, находившихся на рационе с избытком белка.
Содержание аα1 и аα2-глобулинов сыворотки крови в динамике беременности увеличивалось у крыс всех групп. Однако, более значительным их увеличение было у животных, получавших избыточное по углеводам питание. Количество α2-глобулинов увеличилось с 0,1046 ± 0,0072 до 0,1342 ± 0,008, Р < 0,05 у крыс 4-й группы и в контроле соответственно с 0,1046 ± 0,0072 до 0,1282 ± 0,005, Р < 0,05.
Изменения содержания рβ- и уγ-глобулинов носили несущественный характер.
Снижение концентрации белка в сыворотке крови большинство исследователей связывают как с некоторой гидремией, имеющейся у беременных, так и со значительным расходом белка на построение тканей плода.
Снижение содержания альбуминов в сыворотке крови можно, по-видимому, объяснить тем, что с увеличением срока беременности повышается гормональная насыщенность организма и для поддержания гомеостаза возникает необходимость в использовании их в качестве транспортных белков крови, состоящих в основном, из альбуминов (И. А. Мануилова и соавт., 1974).
Отмеченное у животных всех групп некоторое снижение уγ-глобулинов, которые являются носителями бактериальных и вирусных антител, происходит, по-видимому, вследствие передачи их плоду, который в течение определенного времени после рождения не способен их вырабатывать, а несколько более высокий уровень этой фракции у животных 3-й группы, можно полагать, связан с активацией выработки уγ-глобулинов под влиянием избыточного питания.
Изучение показателей углеводного обмена у беременных крыс, находившихся на различных пищевых рационах, показало, что содержание глюкозы в крови животных всех групп во время беременности существенно не изменялось.
Исключение составили животные, получавшие рацион с избытком углеводов; у них содержание сахара в динамике беременности снизилось с (5,52 ± 0,24) до зачатия до (4,86 ± 0,157) ммоль/л во время беременности, Р < 0,01.
Столь парадоксальное явление объясняется тем, что повышение сахара в крови тормозит распад гликогена в печени (М. Ф. Мережинский, 1965; И. Тодоров, 1968). Кроме того, гипергликемия приводит к повышенному вы-делению инсулина клетками р эндокринной части поджелудочной железы.
Содержание пировиноградной и молочной кислот в динамике беременности повышалось у животных контрольной группы и у крыс, получавших рацион с избытком углеводов. Содержание молочной кислоты у животных опытной группы составило до беременности (1,95 ± 0,137) ммоль/л, во время беременности (3,19 ± 0,264) ммоль/л, Р < 0,001; пировиноградной кислоты соответственно (276 ± 15,9) мкмоль/л и (426,0 ± 52,3) мкмоль/л, Р < 0,01.
Увеличение их содержания в крови в течение беременности подтверждено исследованиями ряда авторов (Г. М. Савельева, 1967, 1972; Е. Ф. Штанова, 1970; В. Ф. Григорьев и соавт., 1974; Г. А. Мокрик, 1974, 1980; Andersen и соавт., 1970, и др.) и связано как с напряженностью межуточного обмена в материнском организме (Л. С. Персианинов, 1967; Novy, Edward, 1967; Andersen и соавт., 1970, и др.), так и с метаболизмом плода (Г. М. Савельева, 1966, 1969; Е. Ф. Штанова, 1970; Г. А. Мокрик, 1974; Berg и соавт., 1969; Kornack и соавт., 1969, и др.).
Показатели липидного обмена представлены в табл. 1. Из приведенных в таблице данных видно, что содержание общих липидов и холестерина у всех крыс в динамике беременности значительно увеличивалось, достигая наибольшего уровня у животных, получавших рацион с избытком углеводов (4-я группа).
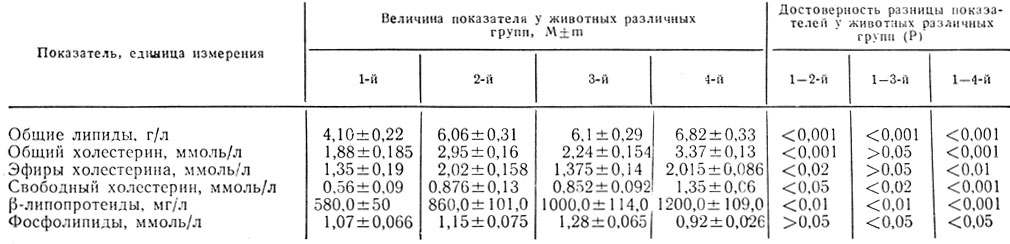
Таблица 1. Показатели жирового обмена у белых крыс, получавших различные пищевые рационы
Содержание p-липопротеидов и фосфолипидов в динамике беременности у животных всех групп повышалось. Исключение составили крысы 4-й группы, у которых отмечена тенденция к его снижению. Обнаруженные к концу беременности изменения липидного обмена соответствуют данным, поученным Т. В. Собко (1975), С. В. Голевым (1977), Х. И. Ялвиром и соавторами (1978), А. А. Закревским (1980), А. Г. Коломийцевой (1981), Л. И. Тутченко
Анализ показателей азотистого обмена и состояния окислительных процессов показал, что количество выводимого азота в суточной моче в динамике беременности у животных, находившихся на рационе с избытком углеводов (4-я группа), уменьшается с (4,5 ± 0,23) до зачатия до (3,7 ± 0,21) ммоль в конце беременности, Р < 0,001, тогда как у крыс, получавших рацион с избытком белка, экскреция общего азота с мочой в динамике беременности возрастает с (4,5 ± 0,23) до (8,4 ± 0,55) ммоль, Р < 0,001.
Выраженная задержка азота у животных, находящихся на рационе с избыточным содержанием углеводов, связана, очевидно, как с влиянием беременности на организм, так и со "сберегающим" белок действием углеводов (А. Е. Браунштейн, М. Г. Крицман, 1967).
Увеличение содержания азота в моче, наблюдаемое у животных, получавших избыток белка, можно объяснить повышением выделения продуктов расщепленного тканевого белка и белка, поступившего с пищей (А М Брейтбург, 1955; Rumsfeld, 1956, и др.).
Исходя из оценки аммиачного коэффициента у крыс, находившихся на различных пищевых рационах, наибольший ацидотический сдвиг наблюдается у животных, содержащихся на рационе с повышенным содержанием углеводов (6,5 ± 0,69), наименьший - у крыс, получавших рацион с избытком белка (3,34 ± 0,49). Установленное избыточное содержание недоокисленных продуктов в организме беременных подтверждают также данные об увеличении ваката кислорода и коэффициента недоокисления мочи у животных, находившихся на рационе с избыточным содержанием углеводов. Уменьшение коэффициента недоокисления мочи к концу беременности у животных, получавших рацион с избытком белка (с 1,1 ± 0,048 до 1,5 ± 0,06, Р < 001), мы связываем с благоприятным влиянием белка на интенсивность течения окислительных процессов. Это подтверждают данные В. В. Суриковой (1967), которая наблюдала значительное уменьшение коэффициента недоокисления мочи при добавлении в пищу людей большого количества белка в виде мяса, а также молочных продуктов.
Для оценки состояния потомства крыс, находившихся во время беременности на качественно различных рационах, проведены исследования 342 крысят от 58 самок, 1-ю группу составляли 162 плода от самок, находившихся на рационе вивария, 2-ю - 90 крысят от самок, получавших рацион с избыточным содержанием белка, и 3-ю - крысят от самок, находившихся на рационе с избыточным содержанием углеводов (табл. 2).
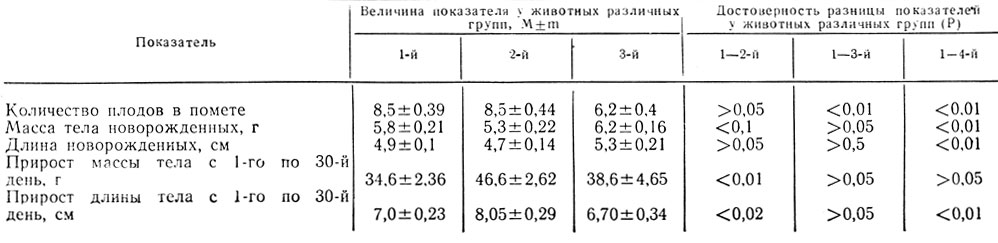
Таблица 2. Количество плодов в помете, показатели физического развития помоства крыс, содержащихся во время беременности на качественно различных рационах
Из табл. 2 следует, что наименьшее число плодов было у крыс 3-й группы; у самок, получавших рацион с избыточным содержанием белка, количество плодов существенно не отличалось от контроля, но было значительно выше, чем в 3-й группе.
Малое количество плодов у самок, находившихся на рационе с избыточным содержанием углеводов, обусловлено повышенной внутриутробной гибелью плодов у этих животных, что можно объяснить снижением содержания белка в рационе до 7,2% по энергетической ценности (в контроле - 18%). Это предположение подтверждается тем, что у самок, находившихся на рационе с избытком белка, уменьшения числа плодов в помете не наблюдалось. Подобные данные были получены Riopelle и соавторами (1975) в опытах на обезьянах. Авторы установили, что при снижении содержания белка в рационе повышается внутриутробная гибель плодов, хотя масса тела детенышей, концентрация общего белка, альбуминов и глобулинов в крови у них не изменяются.
Как следует из табл. 2, масса тела новорожденных крысят у самок 2-й и 3-й групп существенно не отличалась от контроля. В то же время масса тела крысят 3-й группы была выше, чем у крысят 2-й группы.
Длина тела новорожденных крысят 2-й и 3-й групп также существенно не отличалась от контроля. Статистически достоверная разница этого показателя отмечена только между крысятами 2-й и 3-й групп.
Прирост массы и длины тела (с 1-го по 30-й день) был максимальным у крысят 2-й группы.
Следовательно, наибольшей потенцией к росту обладали крысята, родившиеся у самок, получавших во время беременности рацион с избыточным содержанием белка.
Можно высказать предположение, что более высокая масса тела у потомства самок, находившихся во время беременности на рационе с избыточным содержанием углеводов, наблюдается оттого, что углеводы (в виде глюкозы) легко проникают через плаценту путем простой диффузии и используются в последующем для образования гликогена у плода. Кроме того, по мнению Lechtig и соавторов (1975), нарастание массы тела плода при этом идет за счет синтезированного из глюкозы жира. Это согласуется с данными Л. Л. Себискверадзе (1968), которая отмечала достоверное увеличение массы тела плодов при введении глюкозы самкам крыс в период беременности.
Содержание общего белка в сыворотке крови у новорожденных крысят всех групп существенно не отличалось.
Показатели красной крови у новорожденных крысят представлены в табл. 3.
Как следует из табл. 3, у крысят 1-й группы количество эритроцитов существенно не отличалось от контроля, а концентрация гемоглобина в единице объема крови и гематокритное число были выше, чем в контроле.
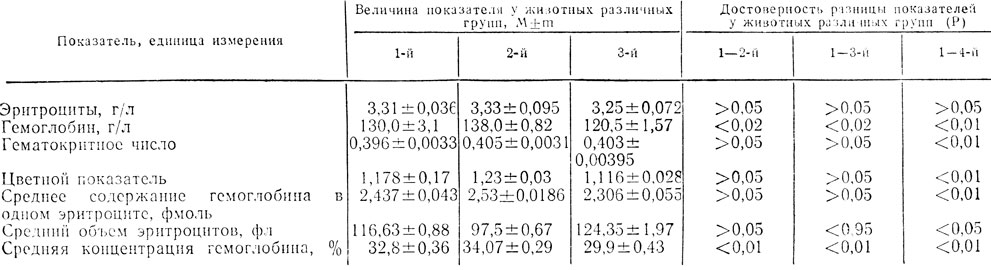
Таблица 3. Показатели красной крови у потомства крыс , находившихся во время беременности на качественно рразличных рационах
Отмечена тенденция к увеличению среднего содержания гемоглобина в одном эритроците по сравнению с контролем, при этом средний объем эритроцита почти не изменился. Наблюдалось также увеличение средней концентрации гемоглобина в одном эритроците, что наряду с изменением других, описанных выше показателей, указывает на увеличение гемоглобинообразования и повышение функциональной активности эритроцитов у крысят, родившихся от самок, получавших рацион с избыточным содержанием белка.
У крысят 3-й группы, по сравнению с контрольными, количество эритроцитов в единице объема крови и гематоритное число существо не отличались от контроля, и в то же время концентрация гемоглобина в единице объема крови была меньше по сравнению с контролем:
Среднее содержание гемоглобина в одном эритроците и средняя его концентрация были меньше, чем в контроле. На основании полученных данных можно предположить, что образование гемоглобина у крысят, рожденных от самок, содержащихся на рационе с повышенным содержанием углеводов, несколько снижено.
Данные о состоянии кроветворения у новорожденных крысят при качественно различном питании самок указывают на то, что развитие плода в значительной мере зависит от характера питания матери.
|
ПОИСК:
|
При копировании материалов активная ссылка обязательна:
http://roghdenierebenka.ru/ 'Беременность, рождение и первые годы жизни ребёнка'